Souvent, pour s’amuser, les hommes d’équipage
Prennent des albatros, vastes oiseaux des mers,
Qui suivent, indolents compagnons de voyage,
Le navire glissant sur les gouffres amers.
(L’Albatros, Baudelaire)
J’écris ces lignes depuis le bout de Bretagne qui a sa place à part dans mon cœur : cette bande qui court de Trégastel à la péninsule de Plougrescant, le long de la Côte de Granit rose et de la Côte des ajoncs. Sur une trentaine de kilomètres, on y voit toujours, au loin, l’archipel des Sept-Îles : l’île aux Moines, l’île Bono, l’île Plate, l’île Malban, le Cerf, les Costans, et surtout l’île Rouzic, à l’est, blanche du guano accumulé par les colonies d’oiseaux de mer qui y résident. C’est là la seule colonie métropolitaine des macareux moines, que j’évoquais ici il y a quelques mois.
C’est d’ailleurs en cherchant à en savoir plus sur ces macareux, sous l’angle de l’écologie évolutive qui a également commencé à trouver sa place ici, que j’ai rencontré cette expression qui a retenu mon attention qui fournira le sujet du jour : le paradoxe des oiseaux de mer (ou seabird paradox dans la littérature), qui nous donnera l’occasion de passer un peu de temps en leur compagnie et à travers quelques concepts d’écologie.
L’expression semble provenir d’un papier de 2008 qui formule le paradoxe ainsi : les oiseaux de mers sont des espèces extrêmement vagiles, capable de se déplacer sur des distances immenses – ce qu’elles font d’ailleurs pour se procurer de la nourriture. Pourtant, beaucoup d’observations ont été faites d’une forte philopatrie chez ces espèces : les individus reviennent se reproduire sur le site de leur naissance (philopatrie natale) ou sur le site de leur première reproduction, limitant par là leur dispersion géographique. Formulé ainsi, le paradoxe porte surtout sur les avantages associés à cette philopatrie qui contrebalanceraient une compétition accrue pour les ressources.
Le paradoxe est parfois aussi formulé sous une autre forme : pourquoi, en dépit de leur vagilité, y a-t-il tant de diversité génétique chez les oiseaux de mer ? Schématiquement, une grande mobilité devrait contribuer à la circulation génétique et à l’homogénéisation des populations, limitant la spéciation. Le lien avec la formulation précédente est que la philopatrie pourrait a priori être un trait limitant cette dispersion, créant des populations suffisamment isolées pour permettre cette spéciation. On va voir que cela n’est en fait pas une bonne explication, mais profitons d’en être ici pour aller regarder du côté des modèles les plus simples reliant structure géographique, mobilité et génétique, pour mieux comprendre le lien entre ces deux versions du paradoxe.
Partons d’abord de la situation la plus simple, celui d’une population de taille 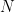
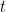


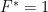
Une première variation sur ce modèle basique est le modèle dit insulaire de Wright : on a désormais à chaque génération une fraction $m$ de nouveaux allèles (tous différents entre eux) injectés dans la population (supposés différents de ceux déjà existants, comme issus d’un réservoir continental infini, amenant par 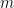
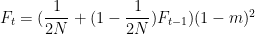
Et, à l’équilibre, 


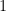
Ce prototype peut être enrichi à volonté, en complexifiant les structures de population et en jouant sur les flux entre différents compartiments de celles-ci. Le cas simple au-dessus permet en tout cas aussi de donner l’exemple le plus basique d’isolement par la distance, en imaginant que le taux
Cette petite digression formelle étant faite, revenons à nos oiseaux de mer… puisqu’en fait l’analyse empirique semble montrer que ce n’est pas ça qui est en jeu. En y regardant de plus près, le paradoxe paraît prendre l’eau de tous côtés. D’une part, il semble y avoir eu des biais dans les mesures historiques conduisant à surestimer la philopatrie des oiseaux de mer – en observant bien plus les individus revenant à leur terre natale que ceux partis nidifier ailleurs.
Même dans des espèces assez philopatriques, comme l’albatros hurleur étudié dans l’article introduisant le paradoxe, on observe un brassage, une homogénéité au niveau génétique qui est bien plus forte que celle observable au niveau démographique – l’homogénéisation des populations par les déplacements des individus n’est donc qu’une partie de l’affaire : le paradoxe apparent au niveau des populations est ici absent au niveau génétique.
Le point vers lequel semblent converger la plupart des études, c’est que ces populations sont en fait loin de l’équilibre migration / dérive génétique, et que l’influence principale sur la diversité génétique (ou son absence) est liée à des facteurs historiques. L’homogénéité génétique des populations d’albatros hurleur viendrait alors d’une extension récente à partir d’une même source peu diverse. A l’inverse, les différences génétiques seraient plutôt expliquées par une fragmentation historique des populations peu reliée à leur distribution actuelle, par exemple durant le dernier maximum glaciaire. On retrouve aussi une trace de cette possibilité dans l’étude sur les macareux : on ne retrouve pas les structures de population visibles dans l’ADN nucléaire quand on regarde l’ADN mitochondrial, dont l’évolution est bien plus rapide, ce qui plaide pour une dynamique encore en cours.
Voilà pour ce petit périple au gré des côtes bretonnes, ces détours biologiques faits, on peut retourner à d’autres divagations poétiques au même endroit, et se concentrer à sentir « que des oiseaux sont ivres d’être parmi l’écume inconnue et les cieux » !